

Original Article: SYSTEMS BIOLOGY of PHOTOSYNTHESIS
Journal: Photobiological Sciences Online
BIOLOGIA DOS SISTEMAS DE FOTOSINÉTICA
Xin-Guang Zhu
Grupo de Biologia de Sistemas de Plantas, Instituto de Biologia Computacional,
Academia Chinesa de Ciências / Max Planck Society, Yue Yang Road,
Shanghai, China, 200031
[email protected]
www.picb.ac.cn/~zhuxinguang
Introdução
Com o esgotamento dos combustíveis fósseis e a crescente demanda por produção de alimentos, os biólogos das plantas pela primeira vez na história estão desempenhando um papel importante na resolução de um grande desafio para a sociedade, ou seja, identificando novas maneiras de aumentar a produtividade das plantas sob um clima global em mudança . Aumentar a eficiência de conversão de energia fotossintética é uma rota importante ainda disponível para que possamos aumentar drasticamente a produtividade das culturas. A combinação de modelos de sistemas com um algoritmo evolutivo fornece um mecanismo eficiente e viável para identificar novas abordagens para engenharia de fotossíntese superior.
Muito do nosso conhecimento sobre a fotossíntese é adquirido através de uma abordagem de redução, onde cada componente individual do sistema é isolado e estudado de forma independente. Através desta abordagem, obtivemos informações biofísicas e bioquímicas detalhadas sobre diferentes componentes da fotossíntese. Ao mesmo tempo, a biotecnologia moderna permite uma regulação rápida ou reduzida de quase todos os genes na fotossíntese. Mesmo com todas essas capacidades e conhecimentos, ainda estamos muito longe de ser capazes de engenharia precisamente do aparelho fotossintético para uma maior taxa fotossintética. Isso não é muito surpreendente, considerando que a eficiência de conversão de energia fotossintética é uma propriedade de sistemas, isto é, é determinada pela interação entre diferentes componentes dos sistemas, não determinada por qualquer componente individual do sistema.
Dado que a fotossíntese é a base da produção de alimentos e energia e do aumento da demanda tanto na nossa sociedade, como podemos engenharia de fotossíntese maior? Em outras palavras, o que é uma limitação da fotossíntese para diferentes plantas em diferentes ambientes? As respostas a estas duas perguntas são a chave para aumentar a produtividade da planta. Responder a estas duas questões é extremamente desafiador. Mais de cem proteínas estão envolvidas na fotossíntese. Mesmo que a manipulação se limite a simplesmente alterar a quantidade de proteína, ao invés de suas propriedades, as permutações potenciais ocorrerão em milhões.
Considerando que várias proteínas podem precisar ser manipuladas ao mesmo tempo, torna-se rapidamente aparente que a regulação excessiva ou a regulação negativa da quantidade de cada proteína e o exame de seus impactos na produtividade são ineficientes. Uma maneira potencial de resolver esse problema é adotar uma abordagem de sistemas. A abordagem de sistemas é desenvolver modelos matemáticos, que podem simular fielmente todos os processos biofísicos e bioquímicos envolvidos na fotossíntese, então, realizar experimentos numéricos para identificar alvos para o engenheiro para maior produtividade.
Utilizarei um modelo de metabolismo de carbono fotossintético para ilustrar a metodologia básica do desenvolvimento de modelos de sistemas e usando um algoritmo evolutivo para identificar a alocação ótima de nitrogênio no metabolismo para uma maior taxa fotossintética de CO2 absorção. Também descreverei brevemente as outras principais aplicações dos modelos de sistemas de fotosíntese, que incluem a ligação de dados genômicos a dados fisiológicos, o estudo de sinais dinâmicos na fotossíntese, o estudo da importância das mudanças no aparelho fotossintético para aumentar a eficiência fotossintética e trabalhar como base para modelos ecofiologicos.
Um dos principais objetivos da pesquisa em biologia vegetal é aumentar a fotossíntese
Durante as últimas quatro décadas, a criação de plantas produziu aumentos notáveis nos rendimentos potenciais de muitas culturas. O rendimento potencial é definido como o rendimento que um genótipo pode atingir sem estresses bióticos e abióticos. Seguindo os princípios de Monteith (1977), o rendimento potencial (Y) de uma cultura e produção primária (Pn) em determinado local é determinado por:
Pn = St.εi. εc/k .... (1a)
Y = η.Pn .... (1b)
Onde St é a integral anual da radiação solar incidente (MJ m-2), εi a eficiência com a qual a radiação é interceptada pela cultura; εc a eficiência com a qual a radiação interceptada é convertida em biomassa; η o índice de colheita ou a eficiência com a qual a biomassa é dividida no produto colhido; e k o conteúdo energético da massa da planta (MJ g-1). St é determinado pelo site e pelo ano. k varia muito pouco entre os órgãos vegetativos, com uma média de 17 MJ / kg, embora os grãos com teor de óleo significativo possam ter maior conteúdo energético (Roberts et al., 1993). Pn é a produtividade primária, ou seja, a biomassa total produzida durante a estação de crescimento. Com base na Equação 1, Y é determinado pelo produto combinado de três eficiências, cada uma descrevendo propriedades fisiológicas amplas da cultura: εi, εc, e η. ε1 é determinado pela velocidade do desenvolvimento e fechamento do dossel, e longevidade, tamanho e arquitetura do dossel. εc é uma função da taxa fotossintética combinada de todas as folhas dentro do dossel, menos perdas respiratórias da cultura (Long et al., 2006).
No contexto da Equação 1, o aumento do rendimento potencial nos últimos 40 anos resultou em grande parte de um aumento de η e εi, enquanto os rendimentos realizados foram melhorados através de uma melhor fertilização e melhoria da proteção das doenças, aumentando εi (revisado: Long et al., 2006). Mas, como podemos aumentar ainda mais? As culturas saudáveis de cultivares modernas em espaçamento otimizado interceptam a maior parte da radiação disponível em sua estação de crescimento, limitando as perspectivas de melhoria εi. O grão nas cultivares modernas de cereais pode representar 60% da biomassa total na colheita (Evan, 1993). O índice de colheita para o grão parece estar se aproximando de um limite superior, uma vez que uma quantidade mínima de biomassa deve permanecer no corpo da planta, para garantir que os nutrientes e reservas vitais possam ser translocados para o grão e para explicar os materiais das paredes celulares que não podem ser degradado. O índice de colheita para culturas de biomassa é ainda maior. E se η e εi estão se aproximando de um limite superior, um aumento adicional no potencial de produção só pode ser alcançado por um aumento de εc, que é determinado pela fotossíntese e respiração.
Então, qual é o máximo teórico? εc, e o que é realizado εc? Uma análise teórica sugere que o máximo εc Para as plantas C3 e C4 são 4,6% e 6%, respectivamente (Figura 1, ver detalhes do cálculo em Zhu et al., 2008).
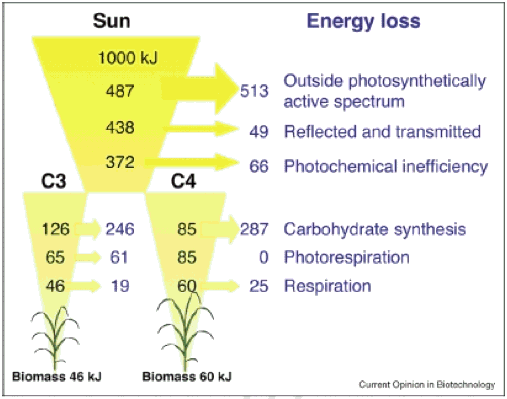
Figura 1. Perdas mínimas de energia calculadas para 1000 kJ de radiação solar incidente, em cada passo discreto do processo fotossintético da planta, desde a intercepção de radiação até a formação de energia química armazenada em biomassa. São consideradas as fotosíntesis C3 e C4 (NADP-Malic Enzyme type). Os cálculos assumem a temperatura da folha de 30°C e uma atmosférica [CO2] de 380 ppm. As setas indicam a quantidade de perdas de energia para diferentes processos (Zhu et al., 2008).
Para as culturas C3, as maiores eficiências a curto prazo são cerca de 0,035, e para C4 cerca de 0,043; comummente alcançado εc no campo é apenas cerca de 1/3 desses valores máximos, sugerindo que há espaço substancial disponível para aumentar ainda mais εc (revisado em Zhu et al., 2008). A evidência direta foi fornecida por plantas cultivadas sob Free Air CO2 Enriquecimento, onde tanto a taxa fotossintética quanto a produtividade da planta são aumentadas (Long et al., 2004).
Como podemos identificar novas formas de engenharia para maior produtividade? Conforme discutido anteriormente, as abordagens experimentais tornam-se ineficientes na identificação de novos objetivos de engenharia. Aqui vou usar o modelo do metabolismo do carbono fotossintético para ilustrar uma abordagem de sistemas para identificar novos alvos para engenharia de maior eficiência de conversão de energia fotossintética.
Abordagem de biologia de sistemas para identificar novas oportunidades de engenharia para maior produtividade
Nesta seção, descreverei brevemente o procedimento básico para o desenvolvimento de um modelo de sistemas e, em seguida, descreva o método de combinação do modelo de sistemas com algoritmos evolutivos para identificar novas metas para o engenheiro para maior produtividade. No final desta seção, vou explicar por que a evolução não selecionou o ótimo aparelho fotossintético.
i) O procedimento básico para desenvolver um modelo de sistemas
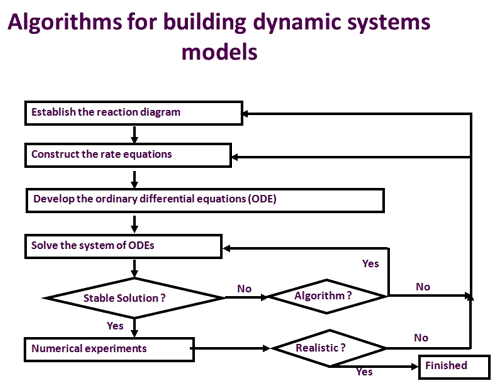
Figura 2. O algoritmo geral para construir um modelo de sistemas dinâmicos. O processo envolvido no desenvolvimento de um modelo de sistemas inclui o estabelecimento do diagrama de reação, a construção das equações de taxa, a construção das equações diferenciais ordinárias, o teste e a validação dos resultados dos modelos com observações experimentais. Este processo itera até que o modelo possa reproduzir fielmente as respostas comumente observadas a perturbações genéticas ou ambientais.
A Figura 2 ilustra o processo utilizado para desenvolver um modelo cinético do processo metabólico. Em primeiro lugar, as reações intimamente envolvidas neste processo precisam ser compiladas para formar um diagrama de reação. Então, a equação de taxa para cada reação envolvida no diagrama é desenvolvida. Um modelo de sistemas que representa este processo metabólico é então construído desenvolvendo uma equação diferencial ordinária (ODE) para cada metabolito envolvido no processo. Cada ODE descreve a taxa de alteração da concentração de metabolitos com o tempo, formada por taxas de formação menos a taxa de consumo deste metabolito. Por exemplo, no metabolismo do carbono fotossintético, o RuBP (Ribulose-1,5-bisphosphate) é formado através da fosforilação Ru5P (Ribulose-5-fosfato) através da reação:
Ru5P+ATP→ ADP + RuBP …. 2
É consumido através da carboxilação RuBP e oxigenação RuBP:
RuBP + CO2 → 2 PGA …. 3
RuBP + O2 → PGA + PGCA …. 4
onde PGA é 3-fosfoglicerato e PGCA é 2-fosfoglicolato. Supondo que a taxa de fosforilação de RuBP seja vrp e as taxas de carboxilação RuBP (Equação 3) e oxigenação (Equação 4) são vc e vo, respectivamente, a taxa de alteração de RuBP com o tempo é expressa como:
d[RuBP]/dt = vrp-vc-vo …. 5
Todas as ODEs formam coletivamente um sistema de equações diferenciais ordinárias, cuja solução corresponde a diferentes estados fisiológicos e dinâmicos do processo metabólico.
Uma vez que um sistema de equações diferenciais comuns foi construído, uma série de experimentos numéricos são necessários para testar e validar o modelo. Normalmente, um modelo de sistemas deve mostrar sua capacidade de ganhar estados estáveis, robustez contra a perturbação externa e a capacidade de prever fenômenos comumente observados. Por exemplo, o modelo do metabolismo do carbono fotossintético atingiu um estado estacionário em cerca de 200 segundos (Zhu et al., 2007). Ele simulou fielmente o comummente medido A-Ci curva, por exemplo, o CO2 absorção versus intercelular CO2 concentração (Ci) curva, para os níveis de oxigênio normal (21%) e baixo (2%). Além disso, após uma drástica perturbação da intercelular O2 concentração, o modelo pode recuperar rapidamente um novo estado estacionário com uma taxa realista de fotossíntese. Finalmente, o modelo simula fisicamente a fotossíntese limitada em condições de baixa taxa de utilização de fosfato de triose (Zhu et al., 2007). Após essas validações, o modelo de metabolismo de carbono fotossintético pode ser usado para identificar novos alvos para engenharia de maior eficiência fotossintética.
ii) Combinando o modelo de sistemas com algoritmo evolutivo para identificar novas metas para o engenheiro para maior produtividade
Na criação tradicional, as cruzes entre diferentes plantas criam variações em certos traços, como o rendimento das culturas. As sementes para pessoas com qualidade de traço superior são selecionadas e usadas para gerar indivíduos para a próxima geração. Após muitas gerações, os traços selecionados são gradualmente aprimorados durante esse processo. Este método levou a um aumento dramático na produtividade das culturas e criou a "revolução verde". Analogamente à seleção de melhores traços ao longo de gerações na criação tradicional, a combinação de modelos de sistemas com um algoritmo evolutivo permite uma evolução simulada do aparelho fotossintético para uma maior taxa fotossintética.
O algoritmo evolutivo imita essencialmente o processo de seleção natural. Na seleção natural real, a pressão de seleção é garantir a sobrevivência e maior fecundidade. Na seleção artificial imitada pelo algoritmo evolutivo, a pressão de seleção é ter uma maior fotosíntese CO2 taxa de absorção com a mesma quantidade de investimento de nitrogênio nas enzimas no metabolismo do carbono fotossintético. Além disso, cada geração tem muitos indivíduos, cada indivíduo representando um conjunto de concentrações de enzimas necessárias para executar o modelo de metabolismo do carbono. Para cada indivíduo, ou conjunto de concentrações de enzimas, existe uma taxa de CO2 absorção. Em cada geração, o indivíduo com maior fotosíntese CO2 a taxa de aceitação será selecionada para "semente" a próxima geração. Para gerar indivíduos para as próximas gerações, o indivíduo selecionado será duplicado para gerar uma população, então os indivíduos dessa população serão "mutados" (isto é, adicionando variações aleatórias às concentrações da enzima). Este processo de seleção e mutação itera, e a taxa de fotosintéticos CO2 A taxa de absorção aumenta gradualmente ao longo das gerações. Este processo termina quando a taxa de fotossíntese não pode ser aumentada com mais gerações (Zhu et al., 2007). A distribuição final de nitrogênio no conjunto de enzimas é considerada a distribuição ótima de nitrogênio (Figura 3).
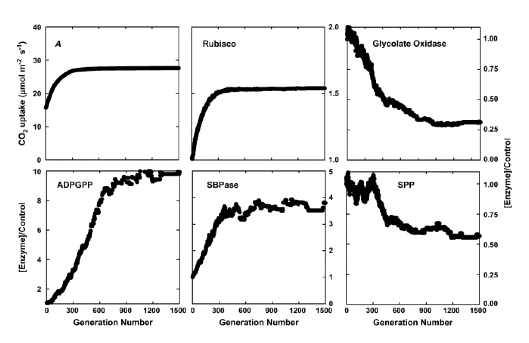
Figura 3. Seleção da partição ideal dos recursos entre as enzimas do metabolismo do carbono para maximizar a taxa fotossintética saturada de luz. A, aumento da taxa fotossintética saturada de luz sobre 1.500 gerações de seleção usando um algoritmo evolutivo. B para F mostram as mudanças nas quantidades de cinco dessas enzimas em relação aos seus valores iniciais ao longo dessas 1.500 gerações. A seleção foi a saturação da luz, e uma Ci de 280 μmol mol-1.
Abreviaturas:
A: fotossintético CO2 taxa de absorção
ADPGPP: ADP pirofosforilase de glicose
SBPase: sedoheptulose-l,7-bisphosphatase
SPP: fosfato de fosfato de sacarose
FBPase: fructose-2,6-bisphosphatase.
Do Zhu et al. (2007).
Uma vez que as distribuições óptimas de nitrogênio são identificadas, podemos usá-la para calcular a demanda de aumento para cada enzima,
Demand[i] = [Enzyme[i]_opt]/ [Enzyme[i]_initial] …. (6)
Onde [Enzima[i]_opt] epresenta a concentração ótima de nitrogênio e [Enzima[i]_initial] representa a concentração inicial de nitrogênio. Se a demanda por uma enzima for superior a 1, a atividade enzimática precisa ser aumentada; Caso contrário, a atividade enzimática precisa ser descartada para nitrogênio livre para a síntese de outras enzimas para obter o melhor CO2 taxa de absorção (Zhu et al., 2007). Usando essa abordagem, mostramos que Rubisco, SBPase e FBPase precisam ser aumentados para uma fotossintética mais alta CO2 taxa (Zhu et al., 2007). Além disso, um aumento na capacidade de dissipação, isto é, a capacidade de consumir o fosfato de triose (PGA, ou fosfato de dihidroxiacetona ou 3-fosfato de gliceraldeído) através da síntese de amido, sacarose ou outras macromoléculas. O nitrogênio necessário para aumentar as atividades dessas enzimas prevê ser reatribuído a partir da via fotorespiratória (Zhu et al., 2007). A SBPase, uma das enzimas identificadas como aumentadas para obter uma maior fotossíntese, foi exagerada no tabaco, o que levou a uma maior fotossíntese e produção de biomassa (Lefebvre et al., 2005), demonstrando que a combinação de algoritmo evolutivo com modelos de sistemas pode identificar possíveis objetivos de engenharia para uma maior fotossíntese.
Outra abordagem para identificar alvos para o engenheiro é a análise do controle metabólico (MCA) através do cálculo do coeficiente de controle do fluxo (Fell, 1997). Tanto o MCA quanto o algoritmo evolutivo fornecem metas de engenharia; No entanto, existem diferenças fundamentais entre essas duas abordagens. O coeficiente de controle de fluxo de uma enzima representa o controle desta enzima em relação a determinado fluxo, assumindo que todos os outros níveis de enzimas não são alterados, isto é, esse coeficiente representa uma visão local do controle dessa enzima sobre o fluxo do sistema. Por outro lado, o algoritmo evolutivo identifica a distribuição ideal de todas as enzimas no metabolismo ao mesmo tempo que altera todas as concentrações da enzima. Portanto, teoricamente, o algoritmo evolutivo pode identificar alvos para maior fotosíntese a partir de uma visão mais global do sistema.
iii) Por que a fotossíntese, após milhões de anos de evolução, não evoluiu uma taxa fotossintética ideal?
A combinação do algoritmo evolutivo com modelos de sistemas fornece um mecanismo viável e eficiente para identificar o ótimo aparelho fotossintético. Uma questão filosófica que as pessoas podem perguntar é: depois de milhões de anos de evolução, por que as plantas não desenvolveram um óptimo aparelho fotossintético? Existem alguns motivos possíveis. Primeiro, a evolução seleciona a sobrevivência e a fecundidade, e não a produtividade. Para garantir a sobrevivência sob condições de estresse, uma certa quantidade de recursos, como nitrogênio, é investida em mecanismos de proteção em detrimento da eficiência fotossintética. Por exemplo, demonstramos que o nitrogênio é sobre investido na via fotorespiratória (Zhu et al., 2007). Este investimento excessivo, no entanto, é crítico para a sobrevivência das plantas sob condições extremamente quentes e preliminares. Embora tais condições sejam raras, talvez uma vez a cada poucos anos, elas podem ser freqüentes o suficiente para eliminar completamente as plantas sem tais mecanismos de proteção (Zhu et al., 2007). Em segundo lugar, a mudança climática global é outro fator. A atmosfera atmosférica CO2 concentração has been around 280 ppm for the past 25 million years (Barnola et al., 1999), before the rapid increase in the CO2 concentração após a Revolução Industrial. O metabolismo do carbono fotossintético atual, por exemplo, maior investimento na via fotorrespiratória, pode representar uma alocação ótima de nitrogênio para a revolução pré-industrial CO2 níveis (Zhu et al., 2007). Nós já demonstraram que uma das enzimas críticas na fotossíntese, Rubisco, é realmente otimizada para uma maior CO2 absorção a uma atmosfera CO2 concentração de 280 ppm (Zhu et al., 2004). Mais dois motivos sutis também podem ser responsáveis pela atual alocação de nitrogênio não ideal no metabolismo do carbono fotossintético. O investimento sub-ótimo da SBPase pode estar relacionado ao papel da SBPase como um ponto de controle no metabolismo do carbono fotossintético; enquanto o excesso de investimento de enzimas na via fotorrespiratória também pode ser devido à relação rigorosa entre a carboxilação RuBP e a oxigenação para uma dada especificidade de Rubisco e condições atmosféricas (Zhu et al., 2007).
Outras aplicações importantes dos modelos de sistemas de fotossíntese
Os modelos de sistemas dinâmicos possuem uma variedade de outras aplicações, além de identificar novas abordagens para o engenheiro para uma maior produtividade. Esses incluem:
Primeiro, Os modelos de sistemas fornecem um link direto de dados genômicos para dados fenotípicos. As insumos do modelo de sistemas são as atividades enzimáticas ou os conteúdos de proteínas, que podem ser obtidos tanto na pesquisa de literatura quanto na proteômica. As saídas dos modelos de sistemas são concentrações de metabolitos, fluxos e parâmetros fisiológicos.
Segundo, modelo de sistemas pode ser usado para estudar o significado adaptativo das mudanças moleculares no aparelho fotossintético para a eficiência fotossintética. Aqui, o aparelho fotossintético é definido como os vários componentes (enzimas, transportadores de elétrons, etc.) e interações entre diferentes componentes na fotossíntese. Como mencionado anteriormente, a combinação de algoritmos evolutivos ou genéticos com modelo de sistemas fornece uma maneira eficiente de identificar o ótimo aparelho fotossintético em condições definidas. Essa diferença entre o ótimo aparelho fotossintético e o aparelho fotossintético existente oferece oportunidades para engenharia de maior produtividade. Essa diferença também sugere o significado de adaptação de diferentes propriedades do aparelho fotossintético.
Terceiro, modelos de sistemas dinâmicos podem ser usados para testar hipóteses sobre mecanismos subjacentes a sinais dinâmicos relacionados à fotossíntese. Utilizamos um modelo de sistemas dos eventos primários em torno do fotosistema II para estudar a base mecanicista da curva de indução de fluorescência registrada após o brilho de um pulso de luz de saturação (isto é, um pulso de luz curto (< 1s) que pode fechar todos os centros de reação de fotossíntese II) em folhas adaptadas a escuras (Zhu et al., 2005).
Quarto, O modelo de fotosíntese de sistemas pode ser usado em modelos de simulação de culturas ecofisiológicas para permitir a ligação direta do genoma ao ecossistema. O design auxiliado por computador do circuito integrado (IC) desempenhou um papel crítico no rápido desenvolvimento de novas tecnologias IC. Espera-se que um modelo capaz de ligar do genoma ao ecossistema diretamente pode servir o mesmo papel. Embora os modelos de ecossistemas agrícolas tenham sido desenvolvidos ao longo do último meio século, eles ainda não podem prever diretamente o desempenho das culturas do genoma de uma planta e seu ambiente de crescimento (Jones et al., 2003), embora haja poucos esforços recentes para usar QTLs (ou seja, trechos de DNA que estão intimamente ligados aos genes subjacentes a uma característica) em modelos de agroecossistemas. (Yin et al., 2004).
Desenvolver um modelo integrado capaz de vincular diretamente o genoma ao ecossistema é extremamente desafiador, uma vez que esse modelo não só precisa descrever mecanicamente processos em vários níveis organizacionais, desde o nível do ecossistema, até o nível do dossel, os processos de nível foliar e celular, mas também tem para descrever mudanças na expressão gênica sob diferentes condições e diferentes estádios de desenvolvimento. Sem dúvida, desenvolver esse modelo levará anos de esforço conjunto para toda a comunidade de modelagem de plantas. Considerando que a fotossíntese é a base do material e energia para o crescimento e desenvolvimento das plantas, um modelo mecanicista detalhado de fotossíntese é um componente crítico de um modelo integrado. A integração do modelo de metabolismo do carbono fotossintético com modelos agro-ecossistemas pode ser o primeiro passo para a realização de um modelo genoma-ecossistema integrado.
Literatura:
Barnola, J.M., Raynaud, D., Lorius, C. & Barkov, N.I. Historical CO2 registro do núcleo de gelo de Vostok. Em Tendências: um compêndio de Dados sobre Mudanças Globais. Centro de análise de informações de dióxido de carbono, Oak Ridge National Laboratory, Departamento de Energia, Oak Ridge, TN, U.S.A. (1999).
Evans L.T. Evolução da cultura, adaptação e rendimento. CUP, Cambridge. (1993)
Fell, D.A. Compreender o controle do metabolismo. Portland Press, Londres (1997).
Jones, J.W., Hoogenboom, G., Porter, C.H., Boote, K.J., Batchelor, W.D., Hunt, L.A., Wilkens, P.W., Singh, U., Gijsman, A.J., Ritchie, J.T. O modelo de sistema de corte DSSAT. European Journal Of Agronomy 18, 235-265 (2003).
Lefebvre, S., Lawson, T., Zakhleniuk, O.V., Lloyd, J.C. & Raines, C.A. O aumento da atividade da sedoheptulose-1,7-bisfosfatase em plantas de tabaco transgênicas estimula a fotossíntese e o crescimento desde o início do desenvolvimento. Plant Physiol. 138, 451-460 (2005).
Long, S.P., Zhu, X.G., Naidu, S.L. & Ort, D.R. A melhoria da fotossíntese pode aumentar os rendimentos das culturas? Plant Cell Environ. 29, 315-330 (2006).
Long S.P., Ainsworth E.A., Rogers A. e Ort D.R. Aumento do dióxido de carbono atmosférico: as plantas enfrentam seu futuro. Análises Anuais de Plant Biology, 55, 591-628 (2004).
Monteith, J.L. Clima e a eficiência da produção de culturas na Grã-Bretanha. Philos. Trans. R. Soc. Londres 281, 277-294 (1977).
Roberts M.J., Long S.P., Tieszen L.L. e Beadle C.L. Medição da biomassa vegetal e produção primária líquida de vegetação herbácea. In: Fotossíntese e produção em um ambiente em mudança: um manual de campo e laboratório (eds, D.O. Hall, J.M.O. Scurlock, H.R. Bolhàr-Nordenkampf, R.C. Leegood e S.P. Long), pp. 1-21. Chapman & Hall, Londres (1993).
Yin, X.Y., Struik, P.C. & Kropff, M.J. Papel da fisiologia das culturas na predição de relacionamentos genético-fenotípico. Trends Plant Sci. 9, 426-432 (2004).
Zhu, X.-G., Portis Jr., A.R. & Long, S.P. A transformação de plantas C3 com Rubisco estrangeiro aumentaria a produtividade? Uma análise computacional extrapolando das propriedades cinéticas para a fotossíntese do dossel. Plant Cell Environ. 27, 155-165 (2004).
Zhu, X.-G. Abordagens computacionais para orientar a melhoria biotecnológica da Eficiência fotossintética das culturas. Ph.D. tese, a Universidade de Illinois. Urbana, IL. (2004).
Zhu, X.-G., Govindjee, Baker, NR, deSturler, E., Ort, DR, Long, SP Chlorophyll a cinética de indução de fluorescência em folhas predita de um modelo descrevendo cada conjunto discreto de energia de excitação e transferência de elétrons associada ao fotosistema II. Planta 223, 114-133 (2005).
Zhu, X.-G., Sturler, E. & Long, S.P. Otimizando a distribuição de recursos entre as enzimas do metabolismo do carbono pode aumentar drasticamente a taxa fotossintética: uma simulação numérica usando um algoritmo evolutivo. Plant Physiol. 145, 513-526 (2007).
Zhu, X.-G., Long, S.P. & Ort, D.R. Qual é a eficiência máxima com que a fotossíntese pode converter energia solar em biomassa? Tendências em biotecnologia 19, 153-159 (2008).